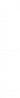Le polyhybridisme est un croisement dans lequel, on étudie plus de deux caractères, chaque caractère étant contrôlé par un seul gène et ces gènes indépendants.
| 1 gène | 2 gènes | 3 gènes | 4 gènes | 5 gènes |
| Mono hybridisme | Di hybridisme | Tri hybridisme | Tetra hybridisme | Penta hybridisme |
| Polyhybridisme | ||||
I- Etude de la transmission de deux caractères (dihybridisme) chez un organisme diploïde.
La transmission de deux couples d’allèles dans le cas du dihybridisme fait intervenir les brassages intra et inter chromosomiques selon la position des deux couples d’allèles sur les chromosomes.
I.1 Brassage inter chromosomique (gènes indépendants).
Les deux gènes gouvernant par exemple deux caractères sont portés par deux paires de chromosomes homologues différentes.
On note A//a B//b deux couples d’allèles théoriques où A et B dominent respectivement a et b.
• Etude d’un cas : Travaux de Morgan sur drosophile melanogaster.
Morgan croise deux mouches de lignées pures : une drosophile à ailes longues et corps gris (souche sauvage) et une drosophile à ailes vestigiales et corps ébène (souche double mutante). A la F1, il obtient 100 % de drosophiles à ailes longues et corps gris (autant de mâles que de femelles).
Puis un deuxième croisement est fait entre une drosophile hybride F1 avec une drosophile à ailes vestigiales et corps ébène. Ils obtiennent :
• 25 % de drosophiles à ailes longues et corps gris ;
• 25 % de drosophiles à ailes vestigiales et corps gris ;
• 25 % de drosophiles à ailes longues et corps ébène ;
• 25 % de drosophiles à ailes vestigiales et corps ébène.
NB : On compte autant de mâles que de femelles pour chaque phénotype.
a) Interprétation des résultats.
À la F1, seuls les allèles « ailes longues » et « corps gris » s’expriment. Ils sont dominants et notés L et G respectivement. Les deux autres allèles « ailes vestigiales » et « corps ébène » sont récessifs et notés v et e.
b) Interprétation génétique.
• Croisement N° 1. • Croisement N° 2 : croisement test ou test cross.
• Croisement N° 2 : croisement test ou test cross.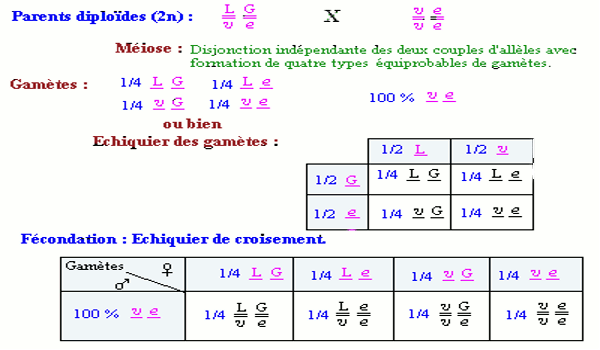 Les proportions \(\frac{1}{4}\), \(\frac{1}{4}\), \(\frac{1}{4}\), \(\frac{1}{4}\) obtenus au test cross caractérisent le dihybridisme avec ségrégation indépendante des couples d’allèles ou gènes indépendants.
Les proportions \(\frac{1}{4}\), \(\frac{1}{4}\), \(\frac{1}{4}\), \(\frac{1}{4}\) obtenus au test cross caractérisent le dihybridisme avec ségrégation indépendante des couples d’allèles ou gènes indépendants.
Troisième loi de Mendel : loi de la ségrégation indépendante des caractères.
Les caractères des différents couples d’allèles mêlés chez les parents de race pure se disjoignent (ou se séparent) indépendamment les uns des autres lors de la formation des gamètes chez l’hybride.
Lors de la gamétogenèse chez l’hybride, il y a séparation indépendante des chromosomes homologues. Chaque cellule haploïde reçoit l’un ou l’autre des deux chromosomes de chaque paire de chromosomes homologues. Dans notre cas précis, il se forme quatre types de gamètes ou quatre lots haploïdes équiprobables : la méiose assure le brassage interchromosomique.
• AUTRE CROISEMENT.
Morgan et ses collaborateurs continuent avec le croisement des hybrides de la F1. ils obtiennent à la deuxième génération :
- 56,3 % de drosophiles à ailes longues et corps gris ;
- 18,7 % de drosophiles à ailes longues et corps ébène ;
- 18,7 % de drosophiles à ailes vestigiales et corps gris ;
- 6,2 % de drosophiles à ailes vestigiales et corps ébène.
• Interprétation génétique de ce croisement.
Il y a disjonction équiprobable des deux couples d’allèles, par conséquent quatre types équiprobables de gamètes dans les proportions \(\frac{1}{4}\) de chaque type. En plus des phénotypes des parents, on obtient deux phénotypes nouveaux (ou recombinés) à savoir « ailes longues, corps ébène » et « ailes vestigiales, corps gris ».
NB : Pour déterminer les gamètes produits par l’hybride, on peut utiliser la méthode suivante :
(\(\frac{1}{2}\) \(\underline L \) + \(\frac{1}{2}\) \(\underline v \)) x (\(\frac{1}{2}\) \(\underline G \) + \(\frac{1}{2}\) \(\underline v \)) = \(\frac{1}{4}\) L \(\underline G \) ; \(\frac{1}{4}\) \(\underline L \) \(\underline e \) ; \(\frac{1}{4}\) \(\underline v \) \(\underline G \) et \(\frac{1}{4}\) \(\underline v \) \(\underline e \).
Comme chaque hybride a produit quatre types de gamètes, on a 16 combinaisons génotypiques possibles lors de la fécondation. On peut les retrouver par un échiquier ou tableau de fécondation.
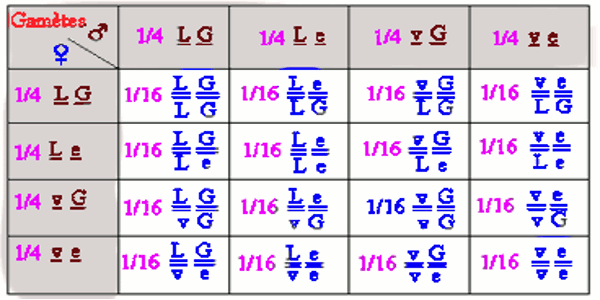

La fécondation amplifie le brassage interchromosomique. La rencontre des gamètes mâles et femelles s’effectuant au hasard, il existe 16 combinaisons diploïdes équiprobables car chaque parent a produit quatre types de gamètes.
Remarque : Tous les individus de la première diagonale sont homozygotes ; ceux de la deuxième diagonale sont tous hétérozygotes.
c) Interprétation chromosomique.
I.2 Brassage intra chromosomique (gènes lies).
Les gènes sont situés sur le même chromosome et ne peuvent pas se séparer de manière indépendante. On dit qu’ils sont liés et constituent un groupe de liaison. C’est ce que les anglo-saxons appellent linkage. On note . \(\frac{{\underline {AB} }}{{ab}}\)
On distingue deux types de liaisons :
• la liaison chromosomique absolue ou totale : les deux couples d’allèles sont transmis ensemble ; les résultats sont identiques à ceux obtenus lors du monohybridisme ;
• la liaison chromosomique partielle ; auquel cas on assiste à des phénomènes de recombinaisons : les deux couples d’allèles peuvent être séparés au cours du brassage intra chromosomique qui a lieu lors de la prophase I de la méiose.
Le brassage intra chromosomique est à l’origine de la diversité des individus et constitue une exception à la troisième loi de Mendel.
I..2.1- Etude d’un exemple.
On croise deux lignées pures de drosophiles : une souche sauvage à ailes longues et yeux rouges et une souche double mutante à ailes vestigiales et yeux pourpres.
A la première génération, on obtient 100 % de drosophiles aux ailes longues et yeux rouges.
• Croisement test N° 1.
On croise une drosophile mâle de la première génération avec une drosophile femelle aux ailes vestigiales et aux yeux pourpres.
Dans la descendance, on note deux phénotypes, tous de type parental, aux proportions égales : 50 % de drosophiles aux ailes longues et yeux rouges et 50 % de drosophiles aux ailes vestigiales et yeux pourpres.
• Croisement test N° 2.
On croise une drosophile femelle de la première génération avec une drosophile mâle aux ailes vestigiales et yeux pourpres.
Dans la descendance, on note la présence de quatre types phénotypiques : a) Interprétation génétique et chromosomique des croisements.
a) Interprétation génétique et chromosomique des croisements.
Les hybrides de la F1 ont tous les phénotypes des parents de la souche sauvage. Les gènes allèles gouvernant les caractères « ailes longues » et « yeux rouges » sont dominants et représentés par L et R respectivement. Par contre, les allèles gouvernant les caractères « ailes vestigiales » et « yeux pourpres » sont récessifs et notés v et p respectivement.
• Croisement test N° 1.
Les résultats obtenus ne sont pas ceux attendus. S’il y avait ségrégation indépendante des gènes allèles, la drosophile double hybride de la F1 devrait produire 4 types de gamètes qui conduiraient à l’apparition de 4 phénotypes avec des fréquences égales : 2 phénotypes parentaux et 2 phénotypes recombinés.
L’apparition de deux phénotypes parentaux exclusivement au test cross résulte du fait que les quatre gènes allèles sont portés par une seule paire de chromosomes homologues. Les mâles de la F1 produisent deux types de gamètes équiprobables \(\underline {LR} \) et \(\underline {vp} \).
On peut expliquer les résultats obtenus en admettant que les deux couples d’allèles sont transmis en bloc : la liaison est totale. Il n’y a donc pas disjonction des couples d’allèles lors de la méiose chez le mâle : c’est une particularité inconnue chez les autres espèces.
• Croisement test N° 2.
Ici, la femelle de la F1 ont formé 4 types de gamètes aux proportions inégales avec un nombre élevé de gamètes de type parental \(\underline {LR} \) et \(\underline {vp} \) et un nombre très faible de gamètes néoformés ou recombinés \(\underline {vR} \) et \(\underline {Lp} \).
Ces résultats sont d’autant plus surprenants que lorsque les gènes sont indépendants, les gamètes apparaissent avec les mêmes chances. On ne peut donc expliquer ce résultat qu’en admettant que lors de la formation des gamètes chez l’hybride diploïde, précisément à la prophase de la première division de la méiose, il s’est produit un crossing-over c'est-à-dire un échange de segments de chromatides homologues. C’est ce qui justifie l’apparition des gamètes nouveaux ou recombinés : la liaison des gènes est dite partielle dans ce cas.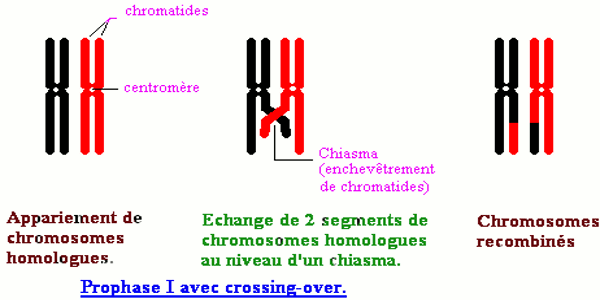 Les proportions des phénotypes parentaux sont supérieures à celles des individus de phénotype nouveau, ce qui caractérise le polyhybridisme avec linkage et crossing-over.
Les proportions des phénotypes parentaux sont supérieures à celles des individus de phénotype nouveau, ce qui caractérise le polyhybridisme avec linkage et crossing-over.
b) Interprétation génétique du croisement N° 1.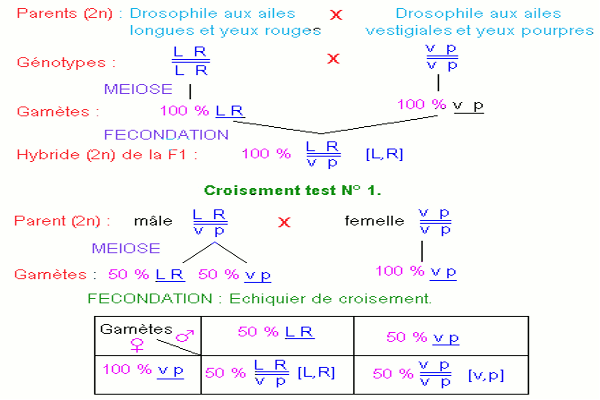 CONCLUSION : Les gènes sont liés de manière absolue sur les chromosomes. Il n’y a pas brassage intra chromosomique et l’information génétique présente dans les gènes est transmise en bloc.
CONCLUSION : Les gènes sont liés de manière absolue sur les chromosomes. Il n’y a pas brassage intra chromosomique et l’information génétique présente dans les gènes est transmise en bloc.
c) Interprétation chromosomique du croisement N° 1.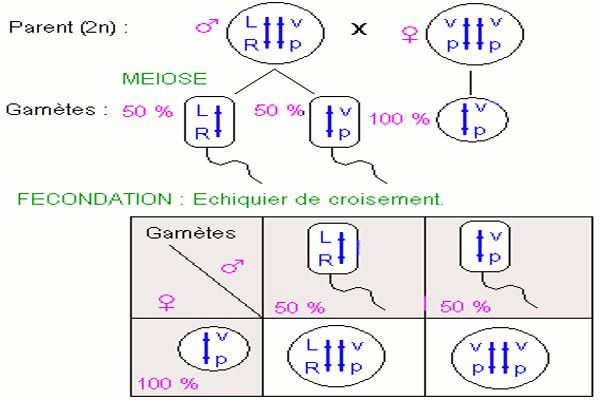 d) Interprétation génétique du croisement test N° 2.
d) Interprétation génétique du croisement test N° 2.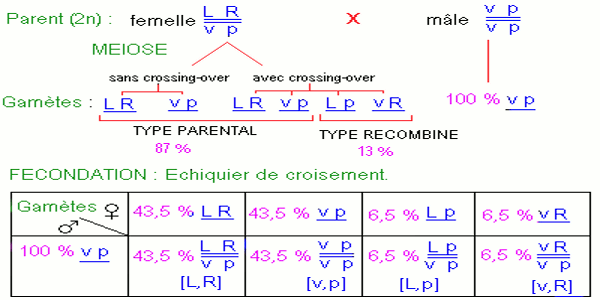 e) Interprétation chromosomique du croisement N° 2.
e) Interprétation chromosomique du croisement N° 2.
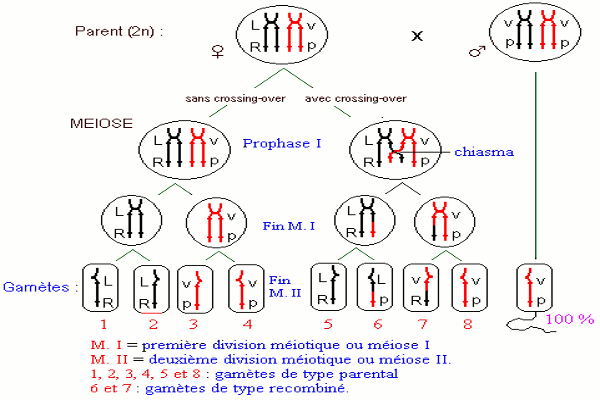
Si on note P le taux de recombinaison, les gamètes recombinés sont produits chacun dans les proportions \(\frac{P}{2}\) tandis que les gamètes non recombinés sont produits dans les proportions \(\frac{{1 - P}}{2}\).
Le taux de recombinaison P se calcule par la formule :
\[\frac{{\sum {{p_N}} }}{{\sum {{p_P}} }} \times 100\]
Avec pN : les phénotypes nouveaux et pP les phénotypes parentaux
I.2.2- Notion de carte factorielle ou génétique.
La probabilité de réalisation d’un crossing-over entre deux gènes donnés dépend de la distance séparant les loci de ces deux gènes. Plus les loci de ces gènes sont distants, plus cette probabilité est grande. La fréquence des gamètes de type recombiné pour deux gènes donnés est directement proportionnelle à la distance entre les loci de ces gènes.
L’unité de distance entre deux loci de deux gènes correspond à 1 % de recombinaison de chromatides soit 1 centimorgan (CM).
L’étude de nombreux croisements concernant différents gènes liés permet de positionner les gènes les uns par rapport aux autres le long du chromosome. La localisation des gènes le long d’un chromosome permet d’établir la carte génétique du chromosome c'est-à-dire une représentation du chromosome indiquant la position relative des loci des gènes les uns par rapport aux autres.
Exemple : P = 13 % soit 13 CM. Conséquence génétique des crossing-over.
Conséquence génétique des crossing-over.
Les échanges de fragments de chromatides entre les chromosomes homologues se font de manière aléatoire le long des chromosomes ; ils sont rares en un endroit précis du chromosome mais plus fréquents à l’échelle du chromosome. Ces échanges intervenant au niveau de chaque paire de chromosomes, créent de nombreuses associations alléliques nouvelles et contribuent à amplifier considérablement la variabilité ou diversité génétique des cellules formées.
Si l’on suppose 100 loci hétérozygotes au niveau d’une paire de chromosomes homologues, \({2^{100}}\) cellules haploïdes différentes peuvent être formées grâce au brassage intra chromosomique
Ainsi, les brassages intra et inter chromosomiques sont les deux phénomènes méiotiques responsables de l’immense diversité génétique des gamètes formés.
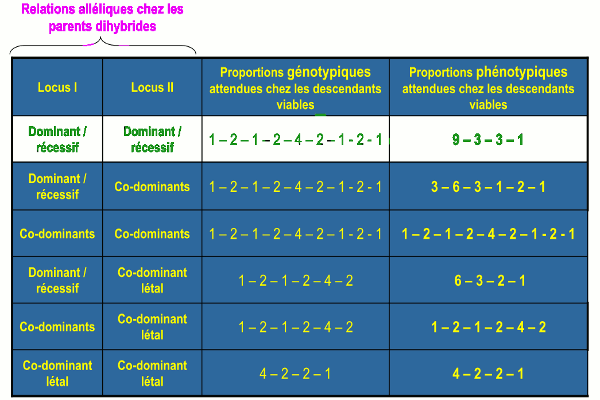
Bilan sur la modification des proportions phénotypiques classiques
Rapports di hybrides modifiés