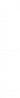Le polyhybridisme est un croisement dans lequel, on étudie plus de deux caractères, chaque caractère étant contrôlé par un seul gène et ces gènes indépendants.
| 1 gène | 2 gènes | 3 gènes | 4 gènes | 5 gènes |
| Mono hybridisme | Di hybridisme | Tri hybridisme | Tetra hybridisme | Penta hybridisme |
| Polyhybridisme | ||||
I- Étude de la transmission de deux caractères (trihybridisme) chez un organisme diploïde.
Nous avons vu qu’un dihybridisme équivaut à l’action simultanée de deux monohybridismes, puisque pour retrouver les proportions de \(\frac{9}{{16}}\), \(\frac{3}{{16}}\), \(\frac{3}{{16}}\) et \(\frac{1}{{16}}\), il suffit de multiplier entre elles les proportions du monohybridisme \(\frac{3}{4}\) et \(\frac{1}{4}\).
Le principe peut également s’appliquer lorsque les croisements font intervenir 3, 4 ou n différences génétiques, à condition que tous les gènes soient situés sur des chromosomes différents.
Les résultats de trihybridisme se prédisent donc aisément :
27 [ABC] 9 [ABc] 9 [AbC] 9 [aBC] 3 [Abc] 3 [aBc] 3 [abC] 1 [abc]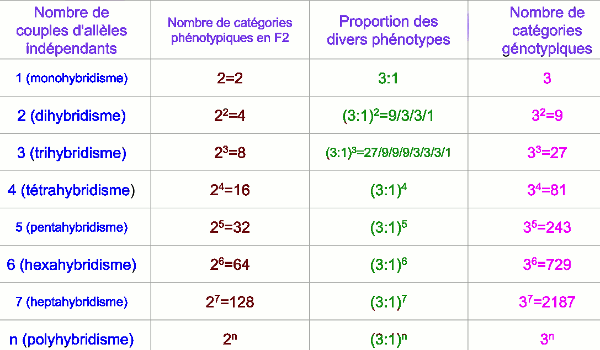 Composition de la descendance de F2 dans un cas de trihybridisme. Il est à noter que les individus caractérisés par la diagonale de ce tableau sont homozygotes pour chacun des caractères.
Composition de la descendance de F2 dans un cas de trihybridisme. Il est à noter que les individus caractérisés par la diagonale de ce tableau sont homozygotes pour chacun des caractères.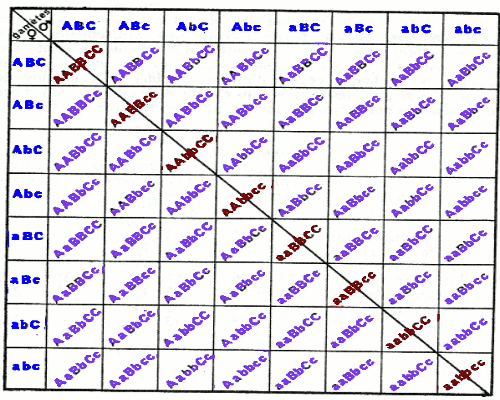 I.1- Cas de trois gènes indépendants.
I.1- Cas de trois gènes indépendants. I.2- Cas des gènes lies.
I.2- Cas des gènes lies.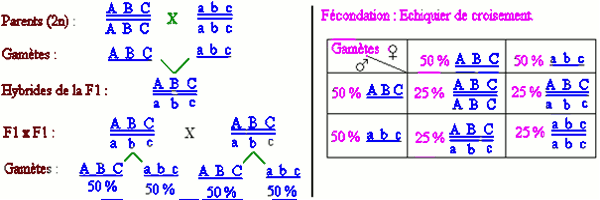 I.2.1- Liaison totale.
I.2.1- Liaison totale.
Application.
On croise des drosophiles mâles au corps jaune (y), aux yeux rouges (ec) et aux ailes tronquées (ct) avec des drosophiles femelles de phénotype sauvage et hétérozygotes pour les trois caractères ([+ + +]).
On obtient 2880 mouches appartenant à huit classes phénotypiques différentes :
- [+ + +] : 1080
- [y ec ct] : 1071 ;
- [+ ec ct] : 66 ;
- [y + +] : 78 ;
- [+ + ct] : 293 ;
- [y ec +] : 283 ;
- [+ ec +] : 6 ;
- [y + ct] : 4.
Etablir la carte génétique indiquant les positions relatives des trois gènes.
Résolution.
Afin de ne pas se tromper dans ce genre de problème, le schéma suivant est nécessaire :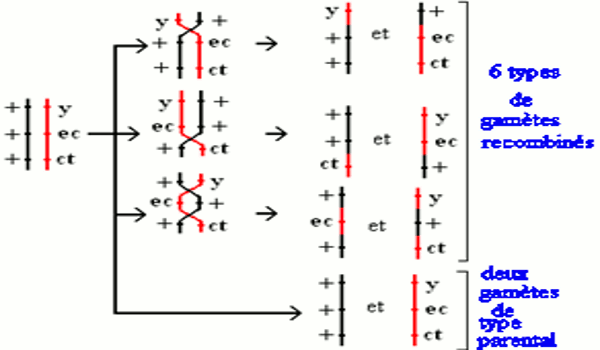 On peut donc avoir quatre types de méiose au cours de la formation des gamètes : des méioses sans crossing-over ; des méioses avec un crossing-over entre les gènes y et ec ; des méioses avec un crossing-over entre les gènes ec et ct, et des méioses avec deux crossing-over : un entre y et ec, l’autre entre ec et ct.
On peut donc avoir quatre types de méiose au cours de la formation des gamètes : des méioses sans crossing-over ; des méioses avec un crossing-over entre les gènes y et ec ; des méioses avec un crossing-over entre les gènes ec et ct, et des méioses avec deux crossing-over : un entre y et ec, l’autre entre ec et ct.
Les taux de recombinaisons sont les suivants :
- Entre y et ec \( = \) \(\frac{{(66 + 78) + (6 + 4)}}{{2880}}\) \( \times 100 = \) \(5,34\% \); ce qui signifie que les deux gènes sont distants de 5,34 CM.
- Entre ec et ct \( = \) \(\frac{{(283 + 293) + (6 + 4)}}{{2880}}\) \( \times 100 = \) \(5,34\% \); ce qui signifie que les deux gènes sont distants de 20,34 CM.
- Entre y et ct \( = \) \(\frac{{(66 + 78) + (283 + 293) + 2(6 + 4)}}{{2880}}\) \( \times 100\) \( = 5,34\% \); donc les deux gènes sont distants de 25,69 CM ; ce qui permet de vérifier les valeurs précédentes.
La carte génétique peut être représentée de la manière suivante :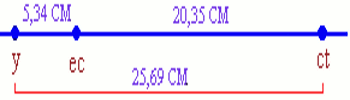 NB : Si l’on envisage la transmission de 4 gènes indépendants ou partiellement liés, on obtient \({2^4}\) soit 16 classes de phénotypes dont deux parentaux et 6 nouveaux \({2^4} - 2\), Le croisement \({F_1} \times {F_1}\) donne \({2^4} \times {2^4}\) \( = {2^8}\) individus uniques.
NB : Si l’on envisage la transmission de 4 gènes indépendants ou partiellement liés, on obtient \({2^4}\) soit 16 classes de phénotypes dont deux parentaux et 6 nouveaux \({2^4} - 2\), Le croisement \({F_1} \times {F_1}\) donne \({2^4} \times {2^4}\) \( = {2^8}\) individus uniques.
Pour n gènes, on a \({2^n} - 2\) phénotypes nouveaux.
La gamétogenèse est caractérisée par un brassage génétique extraordinaire : du fait des brassages intra et inter chromosomiques, l’assortiment génique (combinaison de gènes) contenu dans un gamète n’est que l’un parmi d’innombrables possibles.
En prenant le cas de l’espèce humaine et en supposant que chaque paire de chromosomes comporte en moyenne 100 loci occupés par des couples d’allèles différents, le nombre théorique d’assortiments de gènes serait de \({({2^{100}})^{23}} \) soit \({2^{2300}}\). Autrement dit, un homme pourrait produire \({2^{2300}}\) spermatozoïdes différents et une femme \({2^{2300}}\) ovocytes différents.
Le nombre de types d’œufs représentant la descendance théorique de ce couple serait \({2^{2300}}\) x \({2^{2300}}\) soit \({2^{4600}}\) (ce nombre est de loin supérieur au nombre des atomes présents dans tout l’univers). C’est pourquoi on dit que la fécondation amplifie le brassage génétique.
II- Origine de nouveaux allèles.
Les différents allèles d’un même gène présentent de grandes similitudes, souvent quelques nucléotides seulement diffèrent. Ce constat est à mettre en relation avec la relative instabilité de la molécule d’ADN : c’est par mutation d’un allèle préexistant que se forme un nouvel allèle.
Une mutation est une modification accidentelle de la séquence des nucléotides de l’ADN. La mutation est un phénomène spontané, aléatoire et rare, de l’ordre de 1/1 000 000 (ce sont ses caractéristiques). Toutefois, le risque de mutation peut être considérablement augmenté par certains facteurs de l’environnement dits mutagènes (c'est-à-dire capables de générer des mutations). Ce sont :
- les rayons ionisants (UV, X, rayons émis par les substances radioactives) ;
- les substances chimiques (gaz moutarde, amiante, …).
II.1- Différents types de mutations.
Une mutation peut affecter un seul codon (triplet de nucléotides pouvant coder pour un acide aminé) de l’ADN (mutation ponctuelle) ou au contraire entraîner des remaniements à l’échelle du chromosome (mutation étendues).
II.1.1- Les mutations ponctuelles.
a- Mutations par substitution.
Une seule paire de nucléotides de l’ADN est remplacée par une autre.
Exemple 1:
Avant mutation :
ATG GTG CAC CTG ACT CCT GAG …
TAC CAC GTG GAC TGA GGA CTC…
Après mutation :
ATG GTG CAT CTG ACT CCT GAG …
TAC CAC GTA GAC TGA GGA CTC…
Exemple 2 :
Avant mutation :
GGT AGT TCA AAG GGT…
CCA TCA AGT TTC CCA…
Après mutation :
GGT AGT TAA AAG GGT…
CCA TCA ATT TTC CCA…
a- Mutation par délétion.
Une paire de nucléotide de la molécule d’ADN est perdue. Le « cadre de lecture » des ribosomes est décalée, pouvant faire intervenir précocement un codon stop.
Exemple 3 :
Avant mutation :
GGT TGT TTG CCT CAT TAG C…
CCA ACA AAC GGA GTA ATC G …
Après mutation :
GGT TTT TGC CTC ATT AGC…
CCA AAA ACG GAG TAA TCG …
b- Mutation par addition.
Une nouvelle paire de nucléotides s’insère entre deux nucléotides successifs de la molécule d’ADN. Le « cadre de lecture des ribosomes est également modifié.
Exemple 4 :
Avant mutation :
CCA AAT TCA TAA CTA CTC GG …
GGT TTA AGT ATT GAT GAG CC …
Après mutation :
CCA AAC TTC ATA ACT ACT CGG …
GGT TTG AAG TAT TGA TGA GCC …
Étudier les conséquences de la mutation au niveau du polypeptide à synthétiser.
NB : Les mutations peuvent être classées en fonction des conséquences qu’elles induisent au niveau des polypeptides synthétiser en plusieurs catégories :
- les mutations silencieuses, si la protéine synthétisée ne change pas malgré la modification de l’ADN ;
- les mutations faux sens, si le triplet muté code pour un acide aminé différent ; les conséquences à l’échelle de la cellule et de l’organisme dépendront des modifications structurales et fonctionnelles subies par la protéine ;
- les mutations non sens, si le triplet muté spécifie un arrêt de l’élongation de la chaîne polypeptidique ; la protéine est alors écourtée.
Classer les mutations des exemples 1, 2, 3 et 4.
6.1.2- Les mutations étendues.
a- Les modifications de forme.
Un chromosome peut perdre un fragment (délétion) ou s’enrichir d’un fragment d’un autre chromosome non homologue (translocation). Si le fragment qui se détache se rattache à un chromosome homologue, on parle de duplication.
Un chromosome peut aussi se fragmenter et se recoller après retournement de l’un de ses fragments : c’est une inversion.
Enfin, deux chromosomes non homologues peuvent fusionner en un seul : c’est la fusion.
b- Les modifications de ploïdie.
La ploïdie est le nombre de chromosomes d’une cellule.
En général, les cellules sont diploïdes. Certains organismes présentent un nombre de chromosomes différent du nombre diploïde.
On parle d’aneuploïdie si les chromosomes sont en surnombre ou en infériorité parce qu’une paire de chromosomes homologues ne s’est pas disjointe lors de la méiose.
On parle d’euploïdie si tous les chromosomes sont présents en nombre identique (1, 2, 3 ou plus). Si on a plus de deux jeux de chromosomes par noyau cellulaire, on parle de polyploïdie (triploïdie, tétraploïdie, …). L’apparition de telles formes peut être liée à une gamétogenèse anormale (gamètes diploïdes, …).
Exemple : spermatozoïde 2n + ovocyte n donne une cellule œuf 3n (triploïde).
6.2- Conséquence des mutations.
Quand une mutation affecte un gène, elle produit une version nouvelle de ce gène qui, fréquemment, ne s’exprime pas de façon identique à la version initiale. La cellule mutée réplique son ADN avant de se diviser. La mutation est donc automatiquement transmise aux cellules descendant de la cellule mutée.
Chez un organisme se reproduisant par voie sexuée, une mutation intervenant au niveau d’une de ses cellules ne peut être transmise à la descendance que si cette cellule mutée est à l’origine des cellules sexuelles. Par contre, si la mutation affecte une cellule somatique, elle ne sera pas transmise à la descendance.
6.3- Diversité des allèles.
Un gène est un fragment d’ADN correspondant à un nombre plus ou moins grand de nucléotides.
Dans toute population, de nombreux gènes existent sous plusieurs versions différentes appelées allèles. Chaque version est le résultat d’une mutation intervenue chez un ancêtre et transmise depuis à ses descendants.
Exemple : - le gène ABO qui existe en trois versions différentes A, B et O.
- le système HLA avec 6 gènes très polymorphes : gène DP (6 allèles) ; gène DQ (9 allèles) ; gène DR (46 allèles) ; gène B (52 allèles) ; gène C (11 allèles) et gène A (24 allèles).
Un gène est qualifié de polymorphe si plusieurs allèles sont présents dans la population avec, pour chacun, une fréquence supérieure à 1 % des individus. L’allèle le plus répandu est considéré comme l’allèle normal ; les autres étant des versions mutées.
NB : Plus du tiers des gènes sont polymorphes chez la plupart des êtres vivants. Les mutations jouent donc un rôle important dans l’apparition d’une diversité génétique au sein d’une population.
Conclusion générale.
• Nature du génome.
Un individu possède un patrimoine génétique constitué d’un nombre impressionnant de gènes.
Exemple : le génome humain est formé de 50 à 100 000 gènes.
Dans le génome d’un individu, chaque gène est présent en deux exemplaires (deux allèles). Sauf cas exceptionnel (gènes présents en un seul exemplaire comme ceux porté par la portion spécifique du chromosome Y), le génome est donc un ensemble de couples d’allèles d’un individu.
• Génome et unicité des individus.
Pour chaque gène, un individu hérite de deux allèles parmi les différents allèles possibles présents dans la population ; ces allèles peuvent être identiques (homozygotie) ou non (hétérozygotie).
Si le choix allélique est relativement limité pour certains gènes peu ou pas polymorphes, il est en revanche très grand pour les gènes polymorphes : de nombreux assortiments de deux allèles sont alors possibles. En considérant l’ensemble du génome, on peut dire qu’il y a peu de chances pour que deux individus différents reçoivent le même héritage génétique c'est-à-dire les mêmes combinaisons alléliques. Chaque individu est donc génétiquement unique c'est-à-dire possède un bagage génétique original (exception faite des jumeaux vrais). Ceci est confirmé par les expériences de greffes et les empreintes génétiques.