Objectifs
- Mettre en évidence la diversité génétique au sein d’une population et définir la notion de polymorphisme ;
- Mettre en évidence la variation des fréquences alléliques au sein des populations ;
- Relever l’influence de la sélection naturelle et des migrations sur les variations des fréquences alléliques ;
- Dégager l’importance des variations des fréquences alléliques.
I. Le polymorphisme génétique dans une population.
La génétique des populations est la science qui a pour but de déterminer l’ampleur de la variabilité génétique dans les populations naturelles, d’en expliquer l’origine, le maintien et l’importance évolutive.
Chez les espèces à reproduction sexuée, la diversité génétique est la règle, les individus génétiquement identiques étant des exceptions. Cette diversité des génotypes est à l’origine d’une très grande variation des phénotypes au sein de chaque espèce.
Diverses techniques biochimiques révèlent cette variabilité des phénotypes et génotypes :
• Électrophorèse des protéines ;
• Électrophorèse de l’ADN.
L’électrophorèse est une technique de séparation par un champ électrique des molécules chargées (acides nucléiques, protéines). Les molécules chargées négativement se dirigent vers la cathode et les molécules chargées positivement vers l’anode.
I.1 Notion de population
Le biologiste distingue entre l’individu et l’espèce un groupe d’une importance particulière : la population. Ce terme désigne l’ensemble des individus d’une même espèce qui vivent et se reproduisent entre eux en un lieu assez bien délimité à la même époque.
Étant ainsi plus ou moins isolés sexuellement des populations voisines de la même espèce, on peut considérer que ce groupe d’individus partage un « pool de gènes » propre à la population.
Un pool de gènes est un ensemble de gènes et de leurs différents allèles présents dans une population.
Exemples :
• La population des escargots d’un même champ ;
• Un troupeau de zébus ;
• Les Aborigènes australiens, les Zoulous, les Navajos, …
I.2 Notion de polymorphisme.
Un gène est dit polymorphe si l’allèle le plus commun (normal) a une fréquence inférieure à 0,95. Dans le cas contraire, le gène est dit monomorphe.
Exemples de gènes polymorphes : le gène ABO, les gènes HLA.
Les antigènes d’histocompatibilité du système HLA, Il existe trois principaux gènes codant pour ces protéines : ces gènes dénommés HLA-A, HLA-B. HLA-C, sont tous situes sur le chromosome 6 mais y occupent des locus différents. Pour chacun de ces gènes, on connait de nombreux allèles, on a actuellement identifié dans l’espèce humaine :
• 67 allèles différents pour le gène HLA-A ;
• 149 allèles pour le gène HLA-B ;
• 39 allèles pour le gène HLA-C.
De nombreux mécanismes assurent le maintien du polymorphisme, on peut citer :
• Les mutations spontanées ;
• Les brassages génétiques (inter et intra chromosomique) ;
• Les migrations.
Une population est dite polymorphe si ses individus présentent au moins deux types de phénotypes de façon relativement fréquente. Ici, le terme polymorphisme caractérise ainsi la variabilité des phénotypes existant au sein d’une population.
Le polymorphisme des populations est une conséquence du polymorphisme génique ou polyallélisme.
NB : Les caractères morphologiques du phénotype ne sont pas les seuls pris en compte lorsqu’on parle de polymorphisme. La variabilité des caractères biologiques variés tels que les enzymes, les groupes sanguins, les marqueurs HLA, …) est également étudiée.
II. La variation des fréquences alléliques au sein d’une population
II.1- Calcul des fréquences alléliques
Considérons un seul locus avec deux allèles possibles (A/a), la population peut se répartir en trois génotypes différents (AA, Aa et aa). Si on suppose que l’on a dans la population x individus AA, y individus Aa et z individus aa.
L’effectif total de la population \(N = x + \) \(y + z\); il existe 2N loci au sein de cette population (à cause de la diploïdie).
La fréquence d’un allèle donné est le rapport du nombre de loci occupé par cet allèle et le nombre total de loci existant dans cette population.
• Fréquence de \(A = \) \(\frac{{2x + y + 0z}}{{2N}}\) \( = \) \(\frac{{2x + y}}{{2N}}\) \( = p\)
• Fréquence de \(a = \) \(\frac{{0x + y + 2z}}{{2N}}\) \( = \) \(\frac{{y + 2y}}{{2N}}\) \( = q\)
A partir de ce qui précède, on vérifie que \(p + q = 1\)
La loi de Hardy Weinberg stipule que dans une population d’effectif infini, où les individus se reproduisent au hasard (panmixie), en l’absence des migrations, des mutations et de la sélection naturelle, les fréquences alléliques d’un gène à deux allèles restent constantes d’une génération à l’autre.
II.2 Évaluation le nombre de génotypes possibles au sein d’une population
Si une série allélique comporte m allèles, le nombre de génotypes possibles est de :
\({\left[ {\frac{{m(m + 1)}}{2}} \right]^n}\) ou n le nombre de gènes.
II.3 Influence de la sélection naturelle
La sélection naturelle désigne le phénomène par lequel certains individus de certaines espèces survivent et d'autres sont éliminés, et qui conduit à une évolution et à la conservation des caractéristiques adaptatives de ces espèces.
La viabilité mesure ainsi la probabilité qu’un individu survive depuis la fécondation jusqu’à l’âge reproducteur. Elle représente la valeur adaptative notée w :
• Pour un génotype qui ne laisse pas de descendants, \(w = 0\);
• Pour un génotype ayant une forte aptitude, \(w = 1\).
La valeur sélective, notée s est \(s = 1 - w\).
Sans pression de l’environnement, pratiquement tous les individus d’une même espèce pourraient à la fois muter sans contraintes (avec apparition des caractères phénotypiques correspondants) et survivre. Mais le climat, la végétation, la nourriture ou même la position géographique (près de la mer ou en montagne) avantagent un certain nombre d’individus dont le phénotype se révèle mieux adapté. Par exemple, les animaux montagnards survivent mieux s’ils ont des poils longs, les fleurs si elles attirent les insectes (pour favoriser la pollinisation), etc. Ainsi, la sélection naturelle agit sur les individus par le biais de leur phénotype.
Le résultat direct de cette constatation est que les individus les moins viables se reproduiront beaucoup moins, et que, de fait, leur génotype ne sera pas transmis à la descendance. Ainsi il se produit une sélection du génotype par l’environnement. Les gènes sont ainsi représentés au cours des générations successives en proportion de la valeur sélective de leurs effets phénotypiques (c’est-à-dire des avantages ou des désavantages qu’ils procurent).
Il arrive également que des gènes n’aient aucune influence sur la sélection des individus d’une population donnée, mais puissent néanmoins favoriser son adaptation si, soudain, son milieu de vie change (on parle alors de préadaptation). Une mutation qui ne confère à un individu ni avantage ni désavantage est dite mutation neutre.
a) Étude d’un cas : la drépanocytose chez les humains
Chez certaines populations humaines, l’allèle HbS présente un désavantage à l’état homozygote car il se traduit par une maladie grave de l’hémoglobine. Les homozygotes S//S meurent avant l’âge reproducteur, et ne laissent donc pas de descendance.
Par contre, dans les zones où ne sévit pas le paludisme (ce qui est rare), l’allèle HbA présente un avantage adaptatif. Les homozygotes A//A et les hétérozygotes A//S, souffrant peu ou pas du tout de drépanocytose, sont les plus aptes à se reproduire. Au fil des générations, la sélection naturelle en conservant l’allèle HbA et en éliminant l’allèle HbS accentue les différences entre les fréquences alléliques des deux allèles au sein des populations humaines où sévit la drépanocytose.
b) Notion de polymorphisme équilibre (ou vigueur hybride ou effet hétérosis).
C’est un phénomène observé dans le cas du croisement de deux races pures où les hybrides hétérozygotes présentent un phénotype plus intéressant que celui de chacun des deux parents. On explique ce phénomène par l’augmentation du nombre d’individus hétérozygotes.
Par exemple, dans les zones où sévissent ensemble la drépanocytose et le paludisme, les individus A//A sont victimes de paludisme majoritairement. Les individus S//S sont victimes de drépanocytose.
Les individus A//S sont plus résistants, l’hémolyse rapide des globules rouges empêchant probablement la poursuite du cycle parasitaire. Ils sont donc les plus nombreux dans la population.
Dans ce cas, l’allèle HbS offre un avantage adaptatif.
II.4 Influence des migrations
La migration est le transfert d’animaux provenant d’une population donnée dans une autre population de structure génétique différente.
Les migrations d’individus entre populations différentes, en créant une circulation plus ou moins intense de gènes entre ces populations initialement distinctes, modifient les fréquences alléliques : ainsi, le pool génétique de la population receveuse est enrichie d’une part ; les différences alléliques entre les pools génétiques des deux populations sont réduites. Ces effets sont d’autant plus importants que les deux populations sont plus proches géographiquement (ce qui facilite les échanges génétiques).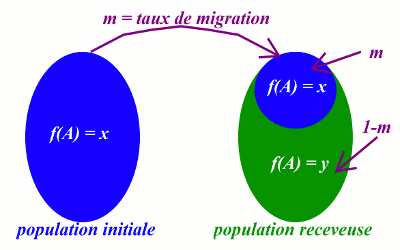 La fréquence de l’allèle A dans la population receveuse après migration est :
La fréquence de l’allèle A dans la population receveuse après migration est :
\(f(A) = m.x\) \( + (1 - m)y\)
Le changement de fréquence de l’allèle A due à l’immigration
\(\Delta P = m\left( {x - y} \right)\)
Avec :
• m = taux de migration c'est-à-dire le rapport du nombre d’immigrants et de l’effectif total de la population receveuse ;
• x = fréquence de l’allèle A dans la proportion des animaux introduits ;
• y = fréquence de l’allèle A dans la population receveuse avant l’immigration.
Remarque : m est petit si on introduit peu d’animaux pour améliorer la population receveuse ; ∆P est par conséquent faible.
II.5 Migrations et effet de fondation
Si les migrations, en mélangeant les pools génétiques, ont tendance à uniformiser ces dernières, elles peuvent aussi créer des populations nouvelles : c’est l’effet de fondation.
En effet, si une migration isole un petit groupe d’individus (sous population) n’ayant emporté qu’une fraction du pool génétique de la population mère, ce groupe peut évoluer indépendamment et constituer une nouvelle population génétiquement originale. Ce mécanisme important est à l’origine de la formation de populations distinctes ; il a lieu lorsque l’effectif d’une espèce est faible et que l’espace géographique pouvant être colonisé est grand. C’est sans doute ainsi que se sont isolées les différentes populations humaines par vagues migratoires successives au cours de l’histoire de notre espèce.
Conclusion
Les variations des fréquences alléliques traduisent les changements dans la composition génétique des populations d’une même espèce. L’évolution est l’accumulation des modifications dans la composition génétique d’une population, dont le résultat est une adaptation accrue à l’environnement. Le processus fondamental de l’évolution réside dans la variation des fréquences alléliques.