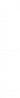III.2 Modes de propagation du message nerveux.
Dans les conditions physiologiques normales, le message nerveux est toujours conduit dans un neurone, des dendrites vers le corps cellulaire puis vers l’axone.
III.2.1 Cas d’une fibre non myélinisée.
L’existence de charges de signes opposées à proximité les unes des autres crée des courants locaux de faible amplitude qui provoquent vers l’avant, du fait de la circulation des électrons du (-) vers le (+), une dépolarisation suffisante pour déclencher un potentiel d’action.
Vers l’arrière, cette même dépolarisation est sans effet du fait de l’existence de la période réfractaire.
III.2.2- Cas d’une fibre myélinisée.
La présence d’une gaine de myéline, isolant électrique de nature lipidique, impose aux courants locaux de s’établir de nœud de Ranvier en nœud de Ranvier. L’influx est donc propagé de manière saltatoire, ce qui explique que la vitesse de conduction des fibres myélinisées soit plus importante que celle des fibres non myélinisées.
NB : Étant recrée de point en point ou de nœud de Ranvier en nœud de Ranvier, le potentiel d’action garde ses caractéristiques (amplitude, durée, surface membranaire) lors de sa conduction (loi du tout ou rien).
III.2.3 Facteurs influençant la vitesse de conduction des potentiels d’action.
o La structure de la fibre nerveuse : une fibre myélinisée conduit l’influx nerveux 50 fois plus vite qu’une fibre amyélinisée.
o Le diamètre ou calibre de la fibre : les fibres les plus grosses sont les plus rapides.
o La température : le froid diminue la vitesse de conduction, la chaleur par contre l’augmente.
Remarque importante: le nerf échappe à la loi du tout ou rien.
 On remplace l’axone par un fragment de nerf dans le dispositif expérimental de la suite 1 Les stimulations efficaces d’intensités croissantes permettent d’obtenir l’enregistrement ci-contre grâce à l’oscilloscope O1.
On remplace l’axone par un fragment de nerf dans le dispositif expérimental de la suite 1 Les stimulations efficaces d’intensités croissantes permettent d’obtenir l’enregistrement ci-contre grâce à l’oscilloscope O1.
Interprétation
Le nerf obéit à la loi de recrutement. Étant constitué de plusieurs fibres nerveuses, une faible intensité de stimulation - supérieure au seuil de stimulation d’une fibre nerveuse – ne touche qu’une ou un petit nombre de fibres
nerveuses. Au fur et à mesure que l’intensité de la stimulation augmente, il y a recrutement d’un nombre de plus en plus grand de fibres nerveuses, ce qui se traduit par l’augmentation de l’amplitude de la dépolarisation. Lorsque toutes les fibres sont recrutées, l’amplitude de la réponse est maximale. Ainsi, au niveau d’un nerf, la dépolarisation observée est une somation des différentes réponses de ses fibres. La réponse du nerf entier à une stimulation efficace est une variation de tension électrique appelée potentiel global.
IV. Mécanismes ioniques à l’origine de l’activité électrique des cellules nerveuses.
L’ultra structure de la membrane plasmique des cellules excitables en général et des neurones en particulier présente principalement deux types de canaux ioniques inégalement répartis :
• des canaux à \({K^ + }\) moins denses (20 canaux /µm2 de surface) ;
• des canaux à \(N{a^ + }\), plus denses (40 canaux / µm2 de surface).
L’ouverture ou la fermeture de ces canaux dépend du potentiel de membrane : on dit que ce sont des canaux ioniques voltage–dépendants.
4.1 Expériences.
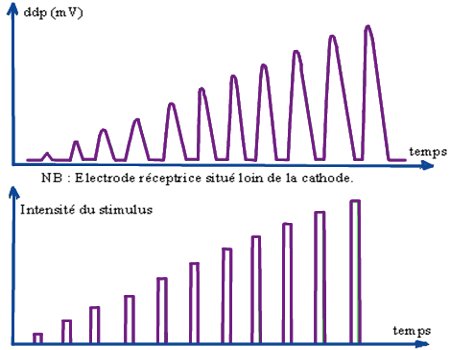
4.1.1- Expérience N° 1.
Une fibre nerveuse est placée dans un milieu contenant de la tétrodotoxine (TTX) qui est une substance qui bloque le fonctionnement des canaux à \(N{a^ + }\).
Une ddp suffisante est imposée à la membrane et on obtient l’enregistrement suivant (voir expérience 1) :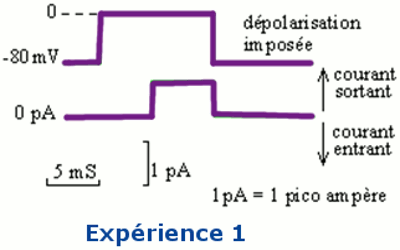
IV.1.2- Expérience N° 2.
Le milieu extracellulaire contient de la tétra-éthyl-ammonium (TEA), qui est une substance bloquant le fonctionnement des canaux de type K+.
On obtient l’enregistrement de l’expérience 2 après dépolarisation.
4.1.3 Expérience N° 3.
Le milieu extracellulaire est non modifié. L’enregistrement de l’expérience N° 3 est obtenu.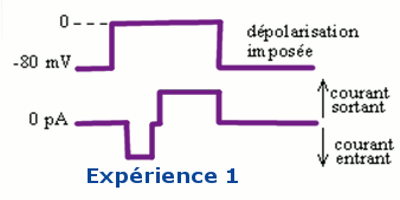
IV.2- Interprétation et conclusion.
Rappel.
Le milieu extracellulaire est riche en \(N{a^ + }\) et le cytoplasme de l’axone est riche en \({K^ + }\).
La TTX bloque les canaux à \(N{a^ + }\). Ainsi le courant sortant enregistré à l’expérience N° 1 est lié à la sortie des ions \({K^ + }\), suite à l’ouverture des canaux à \({K^ + }\) grâce à la dépolarisation suffisante.
La TEA bloque les canaux à \({K^ + }\). Ainsi, le courant entrant enregistré est lié à l’entrée des ions Na+, suite à l’ouverture des canaux à \(N{a^ + }\) grâce à la dépolarisation suffisante.
L’expérience 3 précise enfin la séquence des évènements à l’origine du potentiel d’action :
• une dépolarisation suffisante (quelques dizaines de millivolts) de la membrane nerveuse déclenche l’ouverture des canaux \(N{a^ + }\) voltage-dépendants dans la zone dépolarisée, d’où une entrée des ions \(N{a^ + }\) ;
• la dépolarisation supplémentaire qui en résulte augmente le nombre de canaux Na+ ouverts, et ainsi de suite, la polarisation membranaire s’inversant très vite ;
• les canaux \(N{a^ + }\) ouverts s’inactivent rapidement, interrompant ainsi l’entrée des ions Na+ à leur niveau, alors que des canaux \({K^ + }\) voltage-dépendants s’ouvrent, permettent une sortie d’ions K+ qui repolarise la zone membranaire (et l’hyperpolarise même éventuellement).
Le potentiel de repos est rétabli par le fonctionnement des pompes ioniques à \(N{a^ + }/{K^ + }\).
Les caractéristiques des canaux voltage-dépendants permettent ainsi de comprendre les propriétés de la fibre nerveuse :
o le seuil de dépolarisation nécessaire pour déclencher un potentiel d’action correspond à la valeur de la dépolarisation nécessaire pour ouvrir les canaux \(N{a^ + }\);
o la période réfractaire, qui succède au potentiel d’action et pendant laquelle la membrane est inexcitable, est due à l’inactivation prolongée de ces mêmes canaux \(N{a^ + }\) ;
o la loi du tout ou rien (constante de l’amplitude du potentiel d’action) traduit le fait que cet évènement ne dépend que des concentrations ioniques de part et d’autre de la membrane d’une part, et des propriétés des différents canaux voltage-dépendants impliqués d’autre part. or ces différents paramètres sont stables.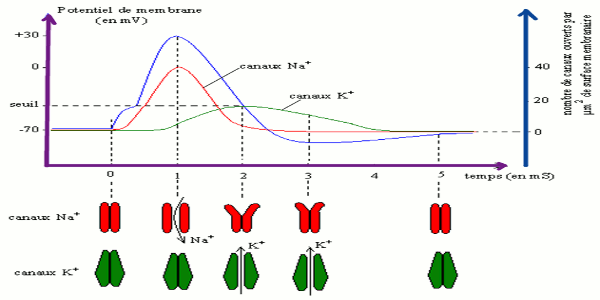 Variations du potentiel de membrane en fonction du nombre de canaux \(N{a^ + }\) et de canaux \({K^ + }\) ouverts.
Variations du potentiel de membrane en fonction du nombre de canaux \(N{a^ + }\) et de canaux \({K^ + }\) ouverts.